
Dòng năng lượng (sinh thái học)
Dòng năng lượng (tiếng Anh: energy flow) là dòng năng lượng truyền qua các sinh vật sống trong hệ sinh thái.[1] Mọi sinh vật sống đều tham gia chuỗi thức ăn, theo vai trò trong chuỗi mà được phân loại thành sinh vật sản xuất, sinh vật tiêu thụ và sinh vật phân giải.[2][3] Mỗi mắt xích hay mỗi loài trong chuỗi thức ăn là một bậc dinh dưỡng.[1] Những mũi tên trong chuỗi thức ăn chỉ hướng của dòng năng lượng một chiều; năng lượng bị mất dưới dạng nhiệt sau khi truyền qua một bậc dinh dưỡng.[2][3] Để thể hiện nhiều thông tin hơn ở mỗi bậc dinh dưỡng, chuỗi thức ăn được tổ chức thành các tháp sinh thái.[1]
Dòng năng lượng một chiều và có thất thoát khi nó truyền qua các bậc của lưới thức ăn là một mô hình chịu sự chi phối của nhiệt động lực học (thuyết trao đổi năng lượng giữa các hệ thống).[4][5] Động lực học dinh dưỡng liên quan đến nhiệt động lực học vì nó liên quan đến vận chuyển và biến đổi năng lượng (có nguồn gốc bên ngoài hệ sinh thái, chủ yếu là từ mặt trời thông qua bức xạ Mặt Trời) và giữa các sinh vật.[1]

Năng lượng và chu trình sinh địa hóa[sửa | sửa mã nguồn]

100×1015 gam carbon/năm được các sinh vật quang hợp cố định, tương đương với 4×1018 kJ/năm = 4×1021 J/năm năng lượng tự do.Hô hấp tế bào là phản ứng ngược, trong đó năng lượng của thực vật được lấy vào, còn carbon dioxide và nước được thải ra. Từ đó carbon dioxide và nước có thể được tái chế trở lại thực vật.
Bước đầu tiên trong quá trình truyền năng lượng trong hệ sinh thái là quang hợp, trong đó nước và carbon dioxide từ không khí được hấp thụ dưới ánh sáng mặt trời, rồi được chuyển đổi thành oxy và glucose.[7] Hô hấp tế bào là phản ứng ngược với quang hợp, trong đó oxy và đường tham gia phản ứng oxy hóa khử và giải phóng năng lượng khi chúng tạo thành carbon dioxide và nước. Carbon dioxide và nước mà hô hấp tạo ra được tái sử dụng trong quang hợp nhờ thực vật.
Thất thoát năng lượng có thể được đo bằng hiệu suất truyền năng lượng (% lượng năng lượng truyền từ bậc dinh dưỡng này sang bậc dinh dưỡng tiếp theo) hoặc sinh khối (% khối lượng vật chất do sinh vật tạo ra truyền từ bậc dinh dưỡng này sang bậc dinh dưỡng tiếp theo).[1] Năng lượng sơ cấp tinh ở sinh vật sản xuất nhìn chung chỉ có 10% chuyển sang bậc tiếp theo (sinh vật tiêu thụ), mỗi bậc dinh dưỡng chỉ truyền được 10% năng lượng sơ cấp tinh lên bậc dinh dưỡng kế tiếp.[1] Hiệu suất sinh thái có giá trị từ 5% đến 20% tùy thuộc vào mức độ hiệu quả của hệ sinh thái đó.[1][8] Hiệu suất sinh thái thấp (10%) chủ yếu do sinh vật hô hấp và năng lượng bị mất dưới dạng nhiệt.[1] Vì hiệu suất sinh thái thấp nên có số cá thể sinh vật tiêu thụ luôn ít hơn so với sinh vật bị tiêu thụ.[1]
Sản xuất sơ cấp[sửa | sửa mã nguồn]
Sinh vật sản xuất là sinh vật tự dưỡng, chủ yếu là sinh vật quang tự dưỡng.[9] Sinh vật sản xuất rất quan trọng vì chúng chuyển đổi năng lượng từ Mặt Trời thành dạng năng lượng hóa học có thể dự trữ và sử dụng là ATP, glucose[1] hay các chất hữu cơ khác và oxy, gọi là năng lượng sơ cấp. Sinh vật sản xuất sử dụng năng lượng do chúng tự dưỡng tạo ra. Nếu sinh vật sản xuất bị động vật ăn cỏ tiêu thụ thì một phần năng lượng từ sinh vật tự dưỡng sẽ được truyền vào động vật ăn cỏ và truyền liên tục như vậy trong chuỗi thức ăn.[1] Glucose trong sinh vật sản xuất được dùng làm thức ăn cho sinh vật tiêu thụ, do đó phải nhờ sinh vật sản xuất mà sinh vật tiêu thụ mới có thể thu năng lượng.[1][7] Sinh vật sản xuất là vi khuẩn tự dưỡng, tảo và thực vật.[1]
Vi khuẩn hóa tự dưỡng thực hiện một quá trình tương tự như quang hợp, nhưng thay vì năng lượng từ Mặt Trời, chúng sử dụng năng lượng dự trữ trong chất vô cơ như hydro sunfide.[10][11] Quá trình này (được gọi là hóa tổng hợp) thường xảy ra sâu trong đại dương tại những miệng phun thủy nhiệt có sẵn nhiệt độ và các hóa chất như hydro, hydro sulfide và metan.[10] Vi khuẩn hóa tự dưỡng có thể sử dụng năng lượng trong các liên kết của hydro sunfide và oxy để chuyển đổi carbon dioxide thành glucose, giải phóng nước và lưu huỳnh trong quá trình này.[11] Sinh vật tiêu thụ vi khuẩn hóa tổng hợp có thể hấp thụ glucose và sử dụng oxy để tiến hành hô hấp tế bào, tương tự như động vật ăn cỏ tiêu thụ sinh vật sản xuất.
Một trong những yếu tố kiểm soát sản xuất sơ cấp là lượng năng lượng đi vào sinh vật sản xuất, có thể được đo đếm bằng hiệu suất sinh thái.[1][12][13] Chỉ 1% năng lượng Mặt Trời đi vào sinh vật sản xuất, phần còn lại phản xạ ra vũ trụ hoặc tán xạ ở khí quyển và mặt đất.[13] Tổng năng suất sơ cấp hay năng suất sơ cấp thô là lượng năng lượng mà sinh vật sản xuất tạo ra được.[13][14] Nhìn chung, 60% năng lượng đi vào cơ thể sinh vật sản xuất được dùng cho hô hấp của chính sinh vật ấy.[12] Năng suất sơ cấp tinh là lượng năng lượng mà thực vật còn lại sau khi trừ đi lượng sử dụng cho hô hấp hoặc rơi rụng...[13] Một yếu tố khác kiểm soát sản xuất sơ cấp là mức độ dinh dưỡng hữu cơ/vô cơ trong nước hoặc đất mà sinh vật sản xuất đang sinh sống.[14]
Sản xuất thứ cấp[sửa | sửa mã nguồn]
Năng lượng thứ cấp là năng lượng mà sinh vật tiêu thụ chuyển đổi thành sinh khối của chính chúng. Những hệ sinh thái khác nhau có mức độ sinh vật tiêu thụ khác nhau, tất cả đều kết thúc bằng một sinh vật tiêu thụ đứng cuối. Hầu hết năng lượng được lưu trữ trong chất hữu cơ của thực vật và khi sinh vật tiêu thụ ăn những thực vật này, chúng sẽ hấp thụ năng lượng này. Năng lượng này trong động vật ăn cỏ và động vật ăn tạp bị động vật ăn thịt tiêu thụ. Ngoài ra còn có một lượng lớn năng lượng trong sản xuất sơ cấp cuối cùng trở thành chất thải hoặc rơi rụng, được gọi là mùn bã. Mùn bã cũng được tiêu thụ bởi sinh vật phân giải, chuỗi thức ăn bắt đầu bằng sinh vật phân giải mùn bã gồm có một lượng lớn vi sinh vật, động vật không xương sống cỡ lớn, động vật không xương sống đáy nhỏ (tiếng Anhː meiobenthos hay meiofauna), nấm và vi khuẩn. Những sinh vật này bị động vật ăn tạp và động vật ăn thịt tiêu thụ, chiếm một lượng lớn năng luọng thứ cấp.[15] Sinh vật tiêu thụ thứ cấp có thể rất khác nhau về hiệu suất tiêu thụ.[16] Hiệu quả của năng lượng truyền cho sinh vật tiêu thụ được ước tính là khoảng 10%.[16] Dòng năng lượng đi qua sinh vật tiêu thụ cao hơn ở môi trường nước so với môi trường trên cạn, chủ yếu do khả năng truyền nhiệt của nước kém hơn đất (nhiệt dung riêng của nước c = 4200 J/kg.K cao hơn nhiều so với nhiệt dung riêng của đất c = 800 J/kg.K), do đó bảo toàn nhiệt độ tốt hơn.
Sản xuất thứ cấp trong môi trường nước
Dị dưỡng đóng góp vào sản xuất thứ cấp, tạo ra năng lượng mà động vật và sinh vật phân hủy sử dụng và nó phụ thuộc vào năng suất sơ cấp tinh.[16] Đa số động vật ăn cỏ và sinh vật phân hủy tiêu thụ carbon từ hai nguồn hữu cơ chính trong hệ sinh thái thủy sinh là nguồn carbon bản địa và ngoại lai.[16] Carbon bản địa đến từ bên trong hệ sinh thái và gồm có thực vật thủy sinh và tảo. Carbon ngoại lai từ ngoài hệ sinh thái chủ yếu là chất hữu cơ chết từ hệ sinh thái trên cạn rơi xuống nước.[16] Trong hệ sinh thái nước chảy, khoảng 66% năng lượng đầu vào thường niên có thể được rửa sạch ở hạ lưu. Lượng còn lại được tiêu thụ và biến mất dưới dạng nhiệt.[17]
Sản xuất thứ cấp trong môi trường trên cạn
Sinh vật tiêu thụ thường có thức ăn ở nhiều bậc dinh dưỡng.[18] Hiệu suất đồng hóa thức ăn được thể hiện bằng lượng thức ăn mà sinh vật tiêu thụ đã ăn, mức độ mà sinh vật tiêu thụ đồng hóa được và mức độ chất thải ra ngoài như phân hoặc nước tiểu. Một phần lớn năng lượng (60 - 70%) hấp thu được từ thức ăn được sinh vật sử dụng để hô hấp, một phần năng lượng khác sẽ chuyển sang sinh khối trong cơ thể sinh vật tiêu thụ.[16] Sinh vật tiêu thụ được chia thành sinh vật tiêu thụ sơ cấp, sinh vật tiêu thụ thứ cấp và sinh vật tiêu thụ bậc cao. Động vật ăn thịt có hiệu suất đồng hóa năng lượng cao hơn nhiều, khoảng 80% và động vật ăn cỏ có hiệu suất thấp hơn nhiều, khoảng 20 đến 50%.[16] Năng lượng trong một hệ thống có thể chịu ảnh hưởng bởi sự di cư/nhập cư của động vật. Sự di chuyển của sinh vật có ý nghĩa trong hệ sinh thái.[17] Mức tiêu thụ năng lượng của động vật ăn cỏ trong hệ sinh thái trên cạn có phạm vi thấp ~3-7%.[17] Sự biến động lượng sản phẩm sơ cấp tinh mà động vật ăn cỏ tiêu thụ nhìn chung là thấp. Điều này trái ngược hoàn toàn với môi trường thủy sinh của hồ và ao, nơi mà động vật ăn cỏ có mức tiêu thụ cao hơn nhiều, khoảng ~33%.[17] Động vật biến nhiệt và động vật đẳng nhiệt có hiệu quả đồng hóa rất khác nhau.[16]
Sinh vật ăn mùn bã[sửa | sửa mã nguồn]
Sinh vật ăn mùn bã tiêu thụ vật chất hữu cơ đang phân hủy và bị động vật ăn thịt tiêu thụ.[16] Năng suất của loài săn mồi có tương quan với năng suất của con mồi, từ đấy khẳng định rằng năng suất sơ cấp trong hệ sinh thái ảnh hưởng đến mọi năng suất kế tiếp.[19]
Mùn bã là một phần lớn chất hữu cơ trong hệ sinh thái. Chất hữu cơ trong rừng ôn đới chủ yếu được hình thành từ thực vật chết, chiếm khoảng 62%.[18]
Trong một hệ sinh thái thủy sinh, lá rơi xuống suối bị ẩm ướt và bắt đầu phân hủy thành chất hữu cơ. Quá trình xảy ra khá nhanh và sẽ thu hút vi sinh vật và động vật không xương sống. Những chiếc lá có thể bị phân hủy thành những mảnh lớn gọi là chất hữu cơ dạng hạt thô (tiếng Anhː coarse particulate organic matter - CPOM).[15] CPOM nhanh chóng bị vi sinh vật tiêu thụ. Động vật không xương sống đáy nhỏ có vai trò cực kỳ quan trọng đối với sản xuất thứ cấp trong các hệ sinh thái nước chảy.[15] Vi sinh vật phân hủy và xâm chiếm CPOM vô cùng quan trọng đối với các loài ăn mùn bã. Loài ăn mùn bã làm cho CPOM trở nên dễ ăn hơn bằng cách giải phóng hợp chất từ các mô và giúp làm mềm chúng.[15] Khi lá thối rữa, nitơ sẽ giảm còn cellulose và lignin trong lá rất khó phân hủy. Sự phân hủy của lá phụ thuộc vào hàm lượng nitơ ban đầu, mùa, loài cây và hàm lượng sinh vật phân giải trong môi trường.
Hiệu suất sinh thái và tính đa dạng loài trong một hệ sinh thái có thể được phân tích thông qua hiệu suất đồng hóa của chúng.[20] Ngoài ra, sản xuất thứ cấp ở hệ sinh thái nước chảy bị ảnh hưởng lớn bởi mùn bã rơi xuống các dòng chảy; sản xuất sinh khối của hệ động vật đáy và độ phong phú giảm thêm 47–50% nếu hệ sinh thái bị mất đi lượng mùn bã này.[19]
Dòng năng lượng qua các hệ sinh thái[sửa | sửa mã nguồn]
Nhiều nghiên cứu đã chứng minh rằng sinh vật sản xuất sơ cấp cố định carbon với tỷ lệ tương tự nhau trong các hệ sinh thái khác nhau.[14] Các cơ chế chi phối dòng năng lượng đến các bậc dinh dưỡng cao hơn sẽ khác nhau giữa các hệ sinh thái. Trong các hệ sinh thái thủy sinh và trên cạn, những mô hình đã được nhận diện có thể giải thích cho sự biến đổi này và được chia làm hai con đường kiểm soát chính: từ trên xuống và từ dưới lên.[21][22] Các cơ chế hoạt động ở mỗi con đường điều chỉnh cấu trúc bậc dinh dưỡng và quần xã trong một hệ sinh thái với những mức độ khác nhau.[23] Con đường kiểm soát từ dưới lên liên quan đến các cơ chế dựa trên chất lượng và tính sẵn có của tài nguyên, kiểm soát năng suất sơ cấp, dòng năng lượng và sinh khối từ bậc dinh dưỡng thấp đến các bậc dinh dưỡng cao hơn.[22] Con đường kiểm soát từ trên xuống liên quan đến các cơ chế dựa trên mức tiêu thụ của sinh vật tiêu thụ.[22][23] Những cơ chế này kiểm soát tỷ lệ vận chuyển năng lượng từ bậc dinh dưỡng này sang bậc dinh dưỡng khác khi động vật ăn cỏ hoặc loài săn mồi ăn các bậc dinh dưỡng thấp hơn.[21]
So sánh dòng năng lượng trong hệ sinh thái thủy sinh và hệ sinh thái trên cạn[sửa | sửa mã nguồn]
Dòng năng lượng sẽ khác nhau trong các hệ sinh thái khác nhau, tạo ra thách thức trong việc xác định khác biệt giữa các loại hệ sinh thái. Theo nghĩa chung, dòng năng lượng là một hàm của năng suất sơ cấp với nhiệt độ, lượng nước và ánh sáng.[24] Ví dụ, ở hệ sinh thái thủy sinh, sản xuất sơ cấp cao hơn thường được tìm thấy ở những dòng sông lớn và hồ cạn hơn là ở hồ sâu và các dòng chảy đầu nguồn nước trong.[24] Ở hệ sinh thái trên cạn, đầm lầy cỏ, đầm lầy cây thân gỗ và rừng mưa nhiệt đới có hiệu suất sản xuất sơ cấp cao nhất, trong khi các hệ sinh thái vùng lãnh nguyên và núi cao có tỷ lệ thấp nhất.[24] Mối quan hệ giữa sản xuất sơ cấp và điều kiện môi trường đã giúp giải thích sự biến đổi trong các loại hệ sinh thái, cho phép các nhà sinh thái học chứng minh rằng dòng năng lượng qua hệ sinh thái thủy sinh hiệu quả hơn so với hệ sinh thái trên cạn do có nhiều cách kiểm soát từ dưới lên và từ trên xuống.[22]
Từ dưới lên[sửa | sửa mã nguồn]
Sức mạnh của các biện pháp kiểm soát từ dưới lên đối với dòng năng lượng được quyết định bởi chất lượng dinh dưỡng, quy mô và tỷ lệ tăng trưởng của sinh vật sản xuất sơ cấp trong hệ sinh thái.[14][21] Chất liệu quang hợp thường giàu nitơ (N) và phosphor (P), bổ sung N và P cho nhu cầu cao của động vật ăn cỏ trong mọi hệ sinh thái.[25] Sản xuất sơ cấp thủy sinh chủ yếu là thực vật phù du đơn bào và nhỏ, chủ yếu bao gồm chất liệu quang hợp, cung cấp nguồn hiệu quả các chất dinh dưỡng này cho động vật ăn cỏ.[21] Ngược lại, thực vật đa bào trên cạn chứa nhiều cấu trúc cellulose hỗ trợ lớn có hàm lượng carbon cao nhưng giá trị dinh dưỡng thấp.[21] Do sự khác biệt về cấu trúc này, sinh vật sản xuất sơ cấp thủy sinh có ít sinh khối trên mỗi mô quang hợp lưu trữ trong hệ sinh thái thủy sinh hơn là trong rừng và đồng cỏ của hệ sinh thái trên cạn.[21] Sinh khối thấp này so với chất liệu quang hợp trong hệ sinh thái thủy sinh cho phép tỷ lệ tuần hoàn hiệu quả hơn so với hệ sinh thái trên cạn.[21] Khi thực vật phù du bị động vật ăn cỏ tiêu thụ, tỷ lệ tăng trưởng và sinh sản của chúng được tăng cường đủ để lấp đầy sinh khối bị mất và cùng với chất lượng đậm đặc dinh dưỡng của chúng, hỗ trợ sản xuất thứ cấp lớn hơn.[21]
Những yếu tố bổ sung tác động đến sản xuất sơ cấp gồm có các đầu vào của N và P, xuất hiện ở mức độ lớn hơn trong hệ sinh thái thủy sinh.[21] Những chất dinh dưỡng này rất quan trọng trong việc kích thích phát triển thực vật và khi được chuyển đến các bậc dinh dưỡng cao hơn sẽ kích thích sinh khối và tỷ lệ phát triển của sinh vật tiêu thụ.[22][24] Nếu một trong hai chất dinh dưỡng này bị thiếu hụt, chúng có thể hạn chế sản xuất sơ cấp tổng thể.[15] Ở các hồ, P có xu hướng là chất dinh dưỡng hạn chế lớn hơn, trong khi cả N và P đều hạn chế sản xuất sơ cấp ở sông.[22] Do những tác động hạn chế này, các chất dinh dưỡng đầu vào có thể làm bớt đi những hạn chế đối với sản lượng sơ cấp ròng của một hệ sinh thái thủy sinh.[23] Chất liệu allochthonous bị cuốn vào hệ sinh thái thủy sinh, mang lại N và P cũng như năng lượng ở dạng phân tử carbon mà sinh vật sản xuất sơ cấp dễ dàng hấp thụ.[15] Đầu vào lớn hơn và nồng độ chất dinh dưỡng tăng lên hỗ trợ tỷ lệ sản xuất sơ cấp ròng cao hơn, từ đó hỗ trợ sản xuất thứ cấp lớn hơn.[25]
Từ trên xuống[sửa | sửa mã nguồn]
Những cơ chế từ trên xuống tạo ra sự kiểm soát lớn hơn lên sinh vật sản xuất sơ cấp thủy sinh do sự luân chuyển sinh vật tiêu thụ trong một lưới thức ăn thủy sinh.[23] Ở sinh vật tiêu thụ, động vật ăn cỏ có thể làm trung gian tác động của các bậc dinh dưỡng bằng cách bắc cầu dòng năng lượng từ sinh vật sản xuất sơ cấp sang động vật ăn thịt ở các bậc dinh dưỡng cao hơn.[26] Trên khắp các hệ sinh thái, có một mối liên hệ nhất quán giữa sự phát triển của động vật ăn cỏ và chất lượng dinh dưỡng của sinh vật sản xuất.[25] Tuy nhiên, ở hệ sinh thái thủy sinh, sinh vật sản xuất sơ cấp bị động vật ăn cỏ tiêu thụ với tỉ lệ cao gấp bốn lần so với các hệ sinh thái trên cạn.[21] Mặc dù đề tài này đang được tranh luận nhiều, nhưng các nhà nghiên cứu cho rằng sự khác biệt trong kiểm soát động vật ăn cỏ là do một số lý thuyết, trong đó sinh vật sản xuất đến tỷ lệ kích cỡ sinh vật tiêu thụ và tính chọn lọc của động vật ăn cỏ.[7]
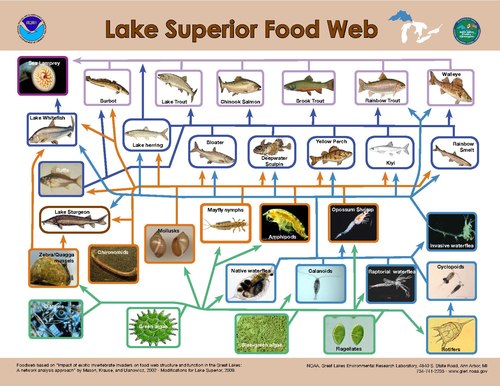
Mô hình kiểm soát từ trên xuống lên sinh vật sản xuất sơ cấp cho thấy rằng việc kiểm soát dòng năng lượng tốt nhất xảy ra khi tỷ lệ kích cỡ của sinh vật tiêu thụ tới sinh vật sản xuất sơ cấp đạt mức cao nhất.[28] Sự phân bố kích cỡ của các sinh vật được tìm thấy trong một bậc dinh dưỡng duy nhất trong các hệ thống thủy sinh hẹp hơn nhiều so với các hệ thống trên cạn.[21] Trên cạn, kích thước của sinh vật tiêu thụ dao động từ nhỏ hơn thực vật mà nó tiêu thụ (chẳng hạn như côn trùng) đến lớn hơn đáng kể (chẳng hạn như động vật móng guốc), trong khi ở các hệ thủy sinh, kích thước cơ thể của sinh vật tiêu dùng trong một bậc dinh dưỡng biến đổi đổi ít hơn nhiều và có mối tương quan chặt chẽ với các vị trí dinh dưỡng.[21] Do đó, sự khác biệt về quy mô giữa sinh vật sản xuất và sinh vật tiêu thụ trong môi trường thủy sinh lớn hơn so với trên cạn, dẫn đến khả năng kiểm soát động vật ăn cỏ mạnh hơn đối với sinh vật sản xuất sơ cấp thủy sinh.[21]
Động vật ăn cỏ có thể kiểm soát số phận của chất hữu cơ khi nó luân chuyển qua lưới thức ăn.[26] Động vật ăn cỏ có xu hướng chọn thực vật giàu dinh dưỡng đồng thời tránh thực vật có cơ chế phòng vệ cấu trúc [21] Giống như cấu trúc hỗ trợ, các cấu trúc phòng vệ bao gồm cellulose nghèo chất dinh dưỡng và có hàm lượng carbon cao.[26] Tiếp cận với các nguồn thực phẩm bổ dưỡng giúp tăng cường trao đổi chất và nhu cầu năng lượng của động vật ăn cỏ, dẫn đến việc loại trừ sinh vật sản xuất sơ cấp đông hơn.[14] Ở hệ sinh thái thủy sinh, thực vật phù du rất giàu dinh dưỡng và thường thiếu cơ chế phòng vệ.[26] Điều này dẫn đến khả năng kiểm soát từ trên xuống lớn hơn vì chất thực vật đã tiêu thụ nhanh chóng được thải lại hệ thống dưới dạng chất thải hữu cơ không bền.[15][26] Ở hệ sinh thái trên cạn, sinh vật sản xuất sơ cấp ít dinh dưỡng hơn và có nhiều khả năng chứa các cấu trúc phòng vệ.[21] Bởi vì động vật ăn cỏ thích thực vật giàu dinh dưỡng và tránh thực vật hoặc các bộ phận của thực vật có cấu trúc phòng vệ, nên một lượng lớn thực vật không được tiêu thụ trong hệ sinh thái.[26] Động vật ăn cỏ tránh chất thực vật chất lượng thấp có thể là do các hệ thống trên cạn thể hiện kiểm soát từ trên xuống yếu hơn so với dòng năng lượng.[21]
Xem thêm[sửa | sửa mã nguồn]
Tham khảo[sửa | sửa mã nguồn]
- ^ a b c d e f g h i j k l m n Lindeman RL (1942). “The Trophic-Dynamic Aspect of Ecology” (PDF). Ecology. 23 (4): 399–417. doi:10.2307/1930126. JSTOR 1930126. Bản gốc (PDF) lưu trữ ngày 29 tháng 3 năm 2017. Truy cập ngày 4 tháng 12 năm 2020.
- ^ a b Briand F, Cohen JE (tháng 11 năm 1987). “Environmental correlates of food chain length”. Science. New York, N.Y. 238 (4829): 956–60. Bibcode:1987Sci...238..956B. doi:10.1126/science.3672136. PMID 3672136.
- ^ a b Vander Zanden MJ, Shuter BJ, Lester N, Rasmussen JB (tháng 10 năm 1999). “Patterns of Food Chain Length in Lakes: A Stable Isotope Study”. The American Naturalist. 154 (4): 406–416. doi:10.1086/303250. PMID 10523487.
- ^ Sharma JP (2009). Environmental studies (ấn bản 3). New Delhi: University Science Press. ISBN 978-81-318-0641-8. OCLC 908431622.
- ^ Van Ness HC (1969). Understanding thermodynamics . New York: Dover Publications, Inc. ISBN 978-1-62198-625-6. OCLC 849744641.
- ^ “Carbon Cycle”. Lưu trữ bản gốc ngày 12 tháng 8 năm 2006.
- ^ a b c d Whitmarsh J, Govindjee (1999). “The photosynthetic process”. Trong Singhal GS, Renger G, Sopory SK, Irrgang KD, Govindjee (biên tập). Concepts in photobiology: photosynthesis and photomorphogenesis. Boston: Kluwer Academic Publishers. tr. 11–51. ISBN 978-0-7923-5519-9..
100×1015 grams of carbon/year fixed by photosynthetic organisms, which is equivalent to 4×1018 kJ/yr = 4×1021 J/yr of free energy stored as reduced carbon.
- ^ Teal JM (1962). “Energy flow in the salt marsh ecosystem of Georgia”. Ecology. 43 (4): 614–624. doi:10.2307/1933451. JSTOR 1933451.
- ^ Morris J, Hartl DL, Knoll AH, Lue R, Michael M (2019). Biology: How Life Works (ấn bản 3). W. H. Freeman. ISBN 978-1319017637.
- ^ a b Kellermann MY, Wegener G, Elvert M, Yoshinaga MY, Lin YS, Holler T, Mollar XP, Knittel K, Hinrichs KU (tháng 11 năm 2012). “Autotrophy as a predominant mode of carbon fixation in anaerobic methane-oxidizing microbial communities”. Proceedings of the National Academy of Sciences of the United States of America. 109 (47): 19321–6. Bibcode:2012PNAS..10919321K. doi:10.1073/pnas.1208795109. PMC 3511159. PMID 23129626.
- ^ a b Cavanaugh CM, Gardiner SL, Jones ML, Jannasch HW, Waterbury JB (tháng 7 năm 1981). “Prokaryotic Cells in the Hydrothermal Vent Tube Worm Riftia pachyptila Jones: Possible Chemoautotrophic Symbionts”. Science. New York, N.Y. 213 (4505): 340–2. Bibcode:1981Sci...213..340C. doi:10.1126/science.213.4505.340. PMID 17819907.
- ^ a b Amthor JS, Baldocchi DD (2001). “Terrestrial higher plant respiration and net primary production”. Terrestrial Global Productivity: 43. doi:10.1016/B978-012505290-0/50004-1. ISBN 9780125052900.
- ^ a b c d Sigman DM, Hain MP (2012). “The biological productivity of the ocean” (PDF). Nature Education Knowledge. 3 (6): 1–6.
- ^ a b c d e Cebrian J (tháng 10 năm 1999). “Patterns in the Fate of Production in Plant Communities”. The American Naturalist. 154 (4): 449–468. doi:10.1086/303244. PMID 10523491.
- ^ a b c d e f g Allan JD, Castillo MM (2007). Stream ecology: structure and function of running waters (ấn bản 2). Dordrecht: Springer. ISBN 978-1-4020-5582-9. OCLC 144222191.
- ^ a b c d e f g h i Smith TM, Smith RL (2015). Elements of ecology (ấn bản 9). Boston. ISBN 978-1-292-07741-3. OCLC 914328590.
- ^ a b c d Fisher SG, Likens GE (tháng 2 năm 1973). “Energy Flow in Bear Brook, New Hampshire: An Integrative Approach to Stream Ecosystem Metabolism”. Ecological Monographs (bằng tiếng Anh). 43 (4): 421–439. doi:10.2307/1942301. JSTOR 1942301.
- ^ a b Hairston Jr NG, Hairston Sr NG (tháng 9 năm 1993). “Cause-Effect Relationships in Energy Flow, Trophic Structure, and Interspecific Interactions”. The American Naturalist. 142 (3): 379–411. doi:10.1086/285546.
- ^ a b Wallace JB, Eggert SL, Meyer JL, Webster JR (tháng 11 năm 1999). “Effects of resource limitation on a detrital‐based ecosystem”. Ecological Monographs. 69 (4): 409–42. doi:10.1890/0012-9615(1999)069[0409:eorloa]2.0.co;2.
- ^ Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, Jouseau C (tháng 10 năm 2006). “Effects of biodiversity on the functioning of trophic groups and ecosystems”. Nature. 443 (7114): 989–92. Bibcode:2006Natur.443..989C. doi:10.1038/nature05202. PMID 17066035.
- ^ a b c d e f g h i j k l m n o p Shurin JB, Gruner DS, Hillebrand H (tháng 1 năm 2006). “All wet or dried up? Real differences between aquatic and terrestrial food webs”. Proceedings. Biological Sciences. 273 (1582): 1–9. doi:10.1098/rspb.2005.3377. PMC 1560001. PMID 16519227.
- ^ a b c d e f La Pierre K, Hanley T (2015). Trophic Ecology: Bottom-Up and Top-Down Interactions Across Aquatic and Terrestrial Systems. Cambridge University Press. tr. 55–85. ISBN 9781316299692.
- ^ a b c d Gruner DS, Smith JE, Seabloom EW, Sandin SA, Ngai JT, Hillebrand H, và đồng nghiệp (tháng 7 năm 2008). “A cross-system synthesis of consumer and nutrient resource control on producer biomass”. Ecology Letters. 11 (7): 740–55. doi:10.1111/j.1461-0248.2008.01192.x. PMID 18445030.
- ^ a b c d Ricklefs RE, Miller GL (2000). Ecology (ấn bản 4). New York: W.H. Freeman & Co. ISBN 0-7167-2829-X. OCLC 40734932.
- ^ a b c Cebrian J, Lartigue J (2004). “Patterns of Herbivory and Decomposition in Aquatic and Terrestrial Ecosystems”. Ecological Monographs. 74 (2): 237–259. doi:10.1890/03-4019.
- ^ a b c d e f Schmitz OJ (tháng 12 năm 2008). “Herbivory from Individuals to Ecosystems”. Annual Review of Ecology, Evolution, and Systematics. 39 (1): 133–152. doi:10.1146/annurev.ecolsys.39.110707.173418.
- ^ Krause AE, Frank KA, Mason DM, Ulanowicz RE, Taylor WW (tháng 11 năm 2003). “Compartments revealed in food-web structure”. Nature. 426 (6964): 282–5. Bibcode:2003Natur.426..282K. doi:10.1038/nature02115. PMID 14628050.
- ^ Shurin JB, Seabloom EW (2005). “The strength of trophic cascades across ecosystems: predictions from allometry and energetics”. Journal of Animal Ecology (bằng tiếng Anh). 74 (6): 1029–1038. doi:10.1111/j.1365-2656.2005.00999.x. ISSN 1365-2656.
Đọc thêm[sửa | sửa mã nguồn]
- Podolinsky S (2004). “Socialism and the Unity of Physical Forces”. Organization & Environment. 17 (1): 61–75. doi:10.1177/1086026603262092.
- Weiner DR (2000). Models of Nature: Ecology, Conservation and Cultural Revolution in Soviet Russia. U.S.: University of Pittsburgh Press.