
Thành viên:Ecology29/Định luật sinh học
Quy luật sinh học (biological rule) hay định luật sinh học hay quy tắc sinh học là một quy luật tổng quát, nguyên tắc hoặc có tính định hướng (quy tắc ngón tay cái) được nghiên cứu, xây dựng, đề xuất để mô tả các khuôn kiểu, mẫu hình quan sát được trong cơ thể sống. Các quy tắc và định luật sinh học thường được phát triển như những cách diễn đạt ngắn gọn có thể áp dụng rộng rãi để giải thích các hiện tượng phức tạp hoặc những quan sát nổi bật về hệ sinh thái và sự phân bố địa lý sinh học của các loài động vật và thực vật trên khắp thế giới, mặc dù chúng đã được đề xuất hoặc mở rộng cho tất cả các loại sinh vật. Nhiều quy luật sinh thái và địa sinh học này được đặt theo tên của các nhà sinh vật học đầu tiên mô tả chúng.
Từ khi khoa học ra đời, các nhà sinh vật học đã tìm cách giải thích những quy luật rõ ràng trong dữ liệu quan sát thực chứng. Trong triết lý sinh học của mình, Aristotle đã suy luận ra các quy tắc chi phối sự khác biệt giữa các động vật bốn chân sống mang thai (theo thuật ngữ hiện đại là động vật có vú có nhau thai trên cạn). Trong số các quy tắc của ông là kích thước cá bố mẹ giảm theo khối lượng cơ thể trưởng thành, trong khi tuổi thọ tăng theo thời kỳ mang thai và theo khối lượng cơ thể, và khả năng sinh sản giảm theo tuổi thọ. Vì vậy, ví dụ, voi có ít bố mẹ hơn chuột, nhưng tuổi thọ và thời kỳ mang thai dài hơn.
Các quy tắc như thế này được tổng kết, đúc kết và phát biểu một cách ngắn gọn từ những thức thu được từ các phép đo khoa học và các thí nghiệm thực chứng ban đầu về thế giới tự nhiên và có thể được sử dụng làm mô hình để dự đoán các quan sát trong tương lai. Trong số các quy tắc sinh học sớm nhất trong thời hiện đại là của Karl Ernst von Baer (từ năm 1828 trở đi) về sự phát triển của phôi, và của Constantin Wilhelm Lambert Gloger về sắc tố động vật vào năm 1833. Có một số hoài nghi giữa các nhà địa lý sinh học về tính hữu ích của các quy tắc chung. Ví dụ, J.C. Briggs, trong cuốn sách Biogeography and Plate Tectonics năm 1987, nhận xét rằng mặc dù các quy tắc của Willi Hennig về cladistics "nói chung là hữu ích" nhưng dù sao thì quy tắc tiến triển của ông là "đáng ngờ".
Định luật di truyền học[sửa | sửa mã nguồn]
NST giới tính và giới tính cơ thể[sửa | sửa mã nguồn]
1. Quy tắc Haldane[sửa | sửa mã nguồn]

Quy tắc Haldane phát biểu rằng nếu trong một loài lai chỉ có một giới là vô sinh thì giới tính đó thường là giới dị giao tử. Giới tính dị giao tử là giới tính có hai nhiễm sắc thể giới tính khác nhau; ở động vật có vú, con đực có nhiễm sắc thể XY. Nó được đặt theo tên của J.B.S. Haldane.[1]
Chọn lọc theo dòng dõi (kin selection)[sửa | sửa mã nguồn]
1. Quy tắc Hamilton[sửa | sửa mã nguồn]
Quy tắc Hamilton là quy tắc trong sinh thái học và sinh học xã hội, công thức toán học do nhà tự nhiên học và nhà di truyền học quần thể người Anh William Donald Hamilton tìm ra, ủng hộ quan điểm cho rằng chọn lọc tự nhiên ủng hộ thành công di truyền chứ không phải thành công sinh sản. Thừa nhận rằng các cá thể có thể truyền bản sao gen của nó cho các thế hệ tương lai thông qua nguồn gốc trực tiếp (nuôi dưỡng con cháu) cũng như gián tiếp bằng cách hỗ trợ sinh sản của những người họ hàng gần (như cháu gái và cháu trai) thông qua hành vi vị tha (là tập tính hy sinh quyền lợi bản thân cá thể thậm chí cả tính mạng vì lợi ích sinh tồn của cả bầy đàn, thông thường trong thế giới động vật, nó mang nghĩa là sự hỗ trợ cho đồng loại).
Định luật sinh học tiến hóa[sửa | sửa mã nguồn]
Địa lý sinh học[sửa | sửa mã nguồn]
1. Quy tắc Foster[sửa | sửa mã nguồn]
.jpg)
Quy tắc Foster, còn được gọi là quy tắc đảo hoặc hiệu ứng đảo, là một quy tắc sinh học, sinh địa lý học trong sinh học tiến hóa phát biểu rằng các thành viên của một loài nhỏ hơn hoặc lớn hơn tùy thuộc vào tài nguyên có sẵn trong môi trường. Ví dụ, người ta biết rằng voi ma mút lùn đã tiến hóa từ voi ma mút thông thường trên các đảo nhỏ. Các con đường tiến hóa tương tự đã được quan sát thấy ở voi, hà mã, trăn Nam Mỹ, lười (chẳng hạn như lười ba ngón lùn), hươu (như hươu Florida) và người.[2][3]
Quy tắc này được J. Bristol Foster phát biểu lần đầu tiên vào năm 1964.[4][5] Trong đó, ông đã so sánh 116 loài trên đảo với các chủng đại lục của chúng. Ông đề xuất rằng một số sinh vật trên đảo phát triển thành phiên bản lớn hơn của chính chúng (insular gigantism) trong khi những loài khác trở thành phiên bản nhỏ hơn của chính chúng (insular dwarfism). Ông đề xuất một lời giải thích đơn giản rằng các sinh vật nhỏ bé hơn sẽ trở thành to lớn hơn khi áp lực săn mồi được giảm bớt do không có một số động vật săn mồi ở đại lục và các sinh vật to lớn hơn trở thành nhỏ bé hơn khi nguồn thức ăn bị hạn chế do hạn chế về diện tích đất.[6]
Ý tưởng đã được Robert MacArthur và Edward O. Wilson mở rộng trong The Theory of Island Biogeography. Năm 1978, Ted J. Case đã công bố một bài báo dài hơn về chủ đề này trên tạp chí Ecology.[7]
Hơn nữa, đã có nghiên cứu vào năm 2019 cho thấy quy tắc Foster có thể áp dụng cho thực vật.[8]
Đồng tiến hoá[sửa | sửa mã nguồn]
Quy tắc Eichler[sửa | sửa mã nguồn]

Quy tắc Eichler cho rằng, loài ký sinh có xu hướng thích nghi với nhiều vật chủ, do đó có vẻ hợp lý khi mong đợi sự đồng biến giữa sự phong phú về thành phần loài của vật chủ và ký sinh trùng của chúng. Wolfdietrich Eichler (1912–1994), nhà động vật học và nhà ký sinh trùng học người Đức[9] là người đầu tiên kết luận về mối quan hệ này vào năm 1942 và sau đó được gọi là Quy tắc Eichler. Đây là một trong ba quy tắc đồng tiến hóa đầu tiên, còn lại là quy tắc Fahrenholz và quy tắc Szidat.[10]
Năm 2012, Vas và các đồng tác giả[11] đã kiểm chứng quy tắc Eichler và kết luận rằng có bằng chứng mạnh mẽ cho thấy mối tương quan đồng biến giữa sự phong phú thành phần loài chim và động vật có vú với sự phong phú chung của vật ký sinh trên chúng.
Quy tắc Fahrenholz[sửa | sửa mã nguồn]
Sự tương ứng chặt chẽ thường thấy giữa các loài ký sinh và vật chủ của chúng đã cho ra quy tắc Fahrenholz, trong đó quy định rằng vật ký sinh và vật chủ của chúng phân hóa đồng bộ. Điều này dẫn đến dự đoán rằng cây phát sinh loài của ký sinh trùng và vật chủ của chúng phải giống hệt nhau về mặt cấu trúc.[12]
Quy tắc Szidat[sửa | sửa mã nguồn]
Quy tắc Szidat cho rằng vật chủ càng nguyên thủy thì ký sinh trùng trong đó càng nguyên thủy.[13]
Khác[sửa | sửa mã nguồn]
Quy tắc Harrison[sửa | sửa mã nguồn]

Quy tắc Harrison là một quan sát (observation) trong sinh học tiến hóa của Launcelot Harrison, phát biểu rằng giữa các loài thân thuộc, kích thước cơ thể vật chủ và ký sinh trùng có xu hướng đồng biến.
Quy tắc Rensch[sửa | sửa mã nguồn]
Quy tắc Rensch là một quy tắc sinh học về tương quan sinh trưởng, liên quan đến mối quan hệ giữa mức độ dị hình kích thước của giới tính và giới tính nào lớn hơn. Giữa các loài trong một dòng dõi, sự dị hình về kích thước tăng khi kích thước cơ thể tăng lên và khi con đực có giới tính lớn hơn. Và giảm khi kích thước cơ thể trung bình tăng lên và khi con cái có giới tính lớn hơn. Quy tắc này được nhà sinh học tiến hóa Bernhard Rensch đề xuất vào năm 1950.[14]
Sau khi kiểm soát các yếu tố gây nhiễu như lịch sử tiến hóa, sự gia tăng kích thước cơ thể trung bình làm cho sự khác biệt về kích thước cơ thể lớn hơn nếu loài có con đực lớn hơn và nhỏ hơn nếu loài có con cái lớn hơn.[15] Một số nghiên cứu đề xuất rằng điều này là do lưỡng tính giới tính (sexual bimaturism), làm cho các đặc điểm của con đực phân hóa nhanh hơn và phát triển trong thời gian dài hơn.[16] Mối tương quan giữa dị hình kích thước của giới tính và kích thước cơ thể được đưa ra giả thuyết là do sự gia tăng cạnh tranh giữa con đực và con đực ở các loài lớn hơn,[17] là kết quả của nguồn tài nguyên môi trường hạn chế, thúc đẩy sự hung hăng giữa các con đực trong việc tiếp cận lãnh thổ sinh sản[18] và các bạn tình giao phối.[15]
Cây phát sinh loài các dòng dõi dường như tuân theo quy tắc này bao gồm các loài linh trưởng, động vật chân vây và động vật guốc chẵn.[19]
Quy tắc này hiếm khi được thử nghiệm trên ký sinh trùng. Một nghiên cứu năm 2019 cho thấy rận ngoại ký sinh Philopteridae và rận Menoponidae tuân theo quy tắc này, trong khi rận ricinid biểu hiện mô hình đảo ngược.[20]
Định luật sinh thái học và cổ sinh vật học[sửa | sửa mã nguồn]
Định luật sinh thái về kích thước và nhiệt độ cơ thể[sửa | sửa mã nguồn]
1. Định luật Kleiber[sửa | sửa mã nguồn]
Định luật Kleiber giải thích rằng nếu động vật càng lớn thì mỗi gram mô trong cơ thể tiêu hao năng lượng càng chậm. Một con chó nặng 10 kg cần ăn khoảng 500 kcal/ngày còn một con voi nặng 1.000 kg (nặng gấp 100 lần) chỉ cần ăn 10.000 kcal (chỉ nhiều gấp 20 lần). Thức ăn ở đại dương rất khan hiếm nên các loài sinh vật này chủ yếu trông chờ vào lượng thức ăn trôi xuống từ tầng nước trên, chúng phải tiến hóa nhằm tăng kích thước cơ thể để tiết kiệm năng lượng tiêu hao.
2. Quy tắc Bergmann[sửa | sửa mã nguồn]
Quy tắc về kích thước cơ thể của Bergmann giải thích rằng, quần thể các loài có kích thước lớn hơn được tìm thấy trong môi trường lạnh hơn, và các loài có kích thước nhỏ hơn được tìm thấy ở các vùng ấm hơn, quy tắc này vẫn còn tranh cãi.
3. Quy tắc Allen[sửa | sửa mã nguồn]

Quy tắc Allen là một quy tắc sinh thái học cho rằng: ở động vật hằng nhiệt thì các bộ phận nhô ra khỏi cơ thể (tai, đuôi, chi) của loài thích nghi với khí hậu lạnh luôn nhỏ hơn của loài tương đồng thích nghi với khí hậu nóng.[21][22][23] Ví dụ: thỏ ôn đới (lạnh) có tai, đuôi và bàn chân nhỏ hơn tai, đuôi và bàn chân của thỏ nhiệt đới (nóng).[24] Theo đó, tỷ số diện tích bề mặt cơ thể với thể tích cơ thể của các loài tương đồng thuộc nhóm động vật hằng nhiệt là khác nhau, biểu hiện sự thích nghi với nhiệt độ môi trường sống.
Quy tắc này được đặt theo tên nhà động vật học người Mỹ Joel Asaph Allen (phát âm IPA: /ăl′ən/) là người đầu tiên phát hiện và mô tả vào năm 1877.[25][26]
4. Quy tắc Cope[sửa | sửa mã nguồn]
Quy tắc Cope là quy tắc sinh học được đặt theo tên của nhà cổ sinh vật học người Mỹ Edward Drinker Cope người đã đưa ra giả thuyết rằng các dòng dõi quần thể có xu hướng tăng trưởng kích thước cơ thể theo thời gian tiến hóa. Nó chưa bao giờ thực sự được Cope phát biểu chính thức mặc dù ông ủng hộ sự xuất hiện của các xu hướng tiến hóa trực hệ. Đôi khi nó còn được gọi là quy tắc Cope-Depéret vì Charles Depéret đã công khai ủng hộ ý tưởng này. Theodor Eimer cũng đã ủng hộ lý thuyết này trong các lần trước đó.
Thuật ngữ "quy tắc Cope" dường như được đặt ra bởi Bernhard Rensch dựa trên thực tế là Depéret đã "bị Cope hóa" trong các tác phẩm của mình. Trong khi quy tắc đã được chứng minh trong nhiều trường hợp, nó không đúng ở tất cả các cấp độ phân loại, hoặc trong tất cả các nhóm. Kích thước cơ thể lớn hơn có liên quan đến việc tăng cường phát triển thể vóc, thể chất, sức sinh sản (sức sinh tồn) vì một số lý do, mặc dù cũng có một số nhược điểm trên cả cá thể và ở cấp độ dòng: các nhóm bao gồm các cá thể lớn hơn dễ bị tuyệt chủng hơn, điều này có thể làm hạn chế kích thước tối đa của sinh vật.
5. Quy tắc Gloger[sửa | sửa mã nguồn]
Quy tắc Gloger là một quy tắc trong sinh thái học nói rằng các loài động vật hằng nhiệt sống trong môi trường nóng ẩm, ví dụ như gần xích đạo, có xu hướng có nhiều sắc tố trên cơ thể hơn các họ hàng của chúng ở vùng lạnh và khô. Quy tắc này được đặt tên sau khi động vật học người Đức Constantin Wilhelm Lambert Gloger, người đầu tiên nhận xét hiện tượng này vào năm 1833 trong một đánh giá về hiệp phương sai giữa khí hậu và màu lông chim.[27] Erwin Stresemann lưu ý rằng ý tưởng này đã được trình bày bởi Pallas trong Zoographia Rosso-Asiatica (1811).[28] Gloger phát hiện ra rằng các loài chim trong môi trường ẩm ướt hơn thì màu lông có xu hướng đậm hơn so với họ hàng của chúng ở vùng khô cằn. Hơn 90% trong số 52 loài chim được nghiên cứu ở Bắc Mỹ phù hợp với quy tắc này.[29]
Định luật cổ sinh vật học[sửa | sửa mã nguồn]
1. Nguyên lý Dollo[sửa | sửa mã nguồn]

Định luật không thể đảo ngược của Dollo được đề xuất vào năm 1893[30] bởi nhà cổ sinh vật học người Bỉ gốc Pháp Louis Dollo nói rằng "một sinh vật không bao giờ trở lại chính xác trạng thái cũ, ngay cả khi nó cảm thấy mình được đặt trong những điều kiện sống tồn tại giống hệt với những điều kiện trước đây mà nó từng sống... nó luôn luôn lưu giữ một số dấu vết của những giai đoạn trung gian mà nó đã trải qua.”[31][32][33]
Định luật sinh lý học và giải phẫu học[sửa | sửa mã nguồn]
1. Định luật Hebb[sửa | sửa mã nguồn]
Định luật Hebb được đề xuất bởi nhà thần kinh học Donald Hebb, phát biểu rằng các kết nối khớp thần kinh được tăng cường khi hai hoặc nhiều tế bào thần kinh được kích hoạt liên tục trong thời gian và không gian. Bằng cách kết hợp việc bắn tế bào tiền ung thư với hoạt động của tế bào sau synap, những thay đổi cấu trúc xảy ra có lợi cho sự xuất hiện của các tổ hợp hoặc mạng lưới thần kinh.[34]
2. Định luật Davis[sửa | sửa mã nguồn]
Định luật Davis được sử dụng trong giải phẫu học và sinh lý học để mô tả cách các mô hình mô mềm tuân theo các yêu cầu áp đặt. Nó tương tự như định luật Wolff áp dụng cho mô xương. Đó là một nguyên tắc sinh lý nói rằng các mô mềm sẽ lành lại theo cách chúng chịu ứng suất về mặt cơ học.
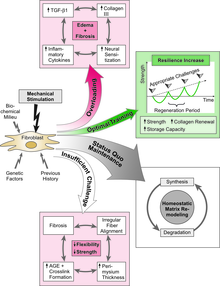
Nó cũng là một ứng dụng thuộc mô hình Mechanostat của Harold Frost ban đầu được phát triển để mô tả phản ứng thích ứng của xương. Tuy nhiên, như chính Harold Frost đã nêu - nó cũng áp dụng cho các mô liên kết dạng sợi, chẳng hạn như dây chằng, gân và mạc cơ (fascia).[35][36] Quy tắc "căng-phì đại" (giãn cơ-tăng kích cỡ) của mô hình đó nêu rõ:"Sự căng ra không liên tục làm cho các mô collagen bị phì đại cho đến khi sự gia tăng sức mạnh làm giảm độ giãn dài của lực căng đến một mức tối thiểu nào đó".[37] Tương tự như hoạt động của các mô xương, phản ứng thích ứng này chỉ xảy ra nếu biến dạng cơ học vượt quá một ngưỡng giá trị nhất định. Harold Frost đề xuất rằng đối với các mô liên kết collagen dày đặc, các ngưỡng giá trị liên quan là khoảng 23 Newton/mm² hoặc độ giãn dài biến dạng 4%.[38]
3. Định luật Wolff[sửa | sửa mã nguồn]
Định luật Wolff sự phát triển bởi nhà giải phẫu học và bác sĩ phẫu thuật người Đức, Julius Wolff (1836–1902) vào thế kỷ 19. Định luật Wolff: dưới tác dụng của lực nép ép, mô xương sẽ phản ứng lại bằng cách tăng sản, phì đại dọc theo đường truyền lực[39] hay cũng được phát biểu rằng xương của một con vật khỏe mạnh sẽ thích ứng với tải trọng mà nó được đặt vào.[40] Như vậy, nơi nào không bị lực tác động thì can xương tiêu đi mà không được thay thế, nhờ đó ống tủy được tái lập và xương được chỉnh trục.
Nếu tải trọng (lực ép) lên một xương cụ thể tăng lên, xương sẽ tự điều chỉnh theo thời gian để trở nên cứng cáp hơn nhằm chống lại loại tải trọng đó.[41][42] Cấu trúc bên trong của bè xương (cũng gọi là thớ cơ hoặc dải cơ) trải qua những thay đổi thích nghi, tiếp theo là những thay đổi thứ cấp ở phần vỏ bên ngoài của xương,[43] có lẽ do đó trở nên dày hơn. Điều ngược lại vẫn đúng: nếu tải trọng lên xương giảm, xương sẽ trở nên kém dàu và yếu hơn do thiếu kích thích cần thiết để tiếp tục tái cấu trúc.[44] Sự giảm mật độ của xương (loãng xương), được biết đến như là sự không chịu được (hiện tượng loãng xương do phần xương trên cán không chịu được - stress shielding) có thể xảy ra do thay khớp háng (hoặc các bộ phận giả khác). Áp lực bình thường lên xương được bảo vệ khỏi xương đó bằng cách đặt trên một bộ phận cấy ghép giả.
Ghi chú[sửa | sửa mã nguồn]
- Quy tắc Fahrenholz
The close correspondence often observed between the taxonomy of parasites and their hosts has led to Fahrenholz's rule, which postulates that parasites and their hosts speciate in synchrony. This leads to the prediction that phylogenetic trees of parasites and their hosts should be topologically identical. We report here a test of this prediction which involves the construction of phylogenetic trees for rodents and their ectoparasites using protein electrophoretic data. We find a high degree of concordance in the branching patterns of the trees which suggests that there is a history of cospeciation in this host-parasite assemblage. In several cases where the branching patterns were identical in the host and parasite phylogenies, the branch lengths were also very similar which, given the assumptions of molecular clock theory, strongly suggests that the speciation of these hosts and ectoparasites was roughly contemporaneous and causally related.
- Đồng tiến hoá
Coevolution is defined as a combination of two processes: co-accommodation between host and parasite with no implication of host or parasite speciation and co-speciation, indicating concomitant host and parasite speciation. Parasite speciation in general is viewed as primarily the result of allopatric speciation processes regardless of host speciation or changes in host type. The observation that co-speciation of hosts and parasites forms a predominant pattern relates to a more general principle of biotic allopatric speciation explicit in the vicariance biogeography model, more than to an assumption that host speciation somehow causes parasite speciation. The close ecological relationship between hosts and parasites, which may be depicted using a variation of the MacArthur-Wilson island biogeography model, explains their spatial proximity at any time during which an isolating event occurs and thus may be necessary in some cases, but is not sufficient to explain parasite speciation, coevolution, or parasite phylogeny.
Tham khảo[sửa | sửa mã nguồn]
- ^ Turelli, M.; Orr, H. Allen (tháng 5 năm 1995). “The Dominance Theory of Haldane's Rule”. Genetics. 140 (1): 389–402. doi:10.1093/genetics/140.1.389. PMC 1206564. PMID 7635302.
- ^ Juan Luis Arsuaga, Andy Klatt, The Neanderthal's Necklace: In Search of the First Thinkers, Thunder's Mouth Press, 2004, ISBN 1-56858-303-6, ISBN 978-1-56858-303-7, p. 199.
- ^ Jean-Baptiste de Panafieu, Patrick Gries, Evolution, Seven Stories Press, 2007, ISBN 1-58322-784-9, ISBN 978-1-58322-784-8, p 42.
- ^ Foster, J. B. (1964). “The evolution of mammals on islands”. Nature. 202 (4929): 234–235. Bibcode:1964Natur.202..234F. doi:10.1038/202234a0.
- ^ Foster J. B. (1965) The evolution of the mammals of the Queen Charlotte Islands, British Columbia. Occasional Papers of the British Columbia Provincial Museum, 14, 1–130.
- ^ Whittaker, R. J. (1998). Island biogeography: ecology, evolution, and conservation. Oxford University Press, UK. tr. 73–75. ISBN 978-0-19-850020-9.
- ^ Case, T. J. (1978). “A general explanation for insular body size trends in terrestrial vertebrates”. Ecology. 59 (1): 1–18. doi:10.2307/1936628.
- ^ Biddick, M.; Hendriks, A.; Burns, K. C. (ngày 19 tháng 8 năm 2019). “Plants obey (and disobey) the island rule”. Proceedings of the National Academy of Sciences (bằng tiếng Anh): 201907424. doi:10.1073/pnas.1907424116. ISSN 0027-8424.
- ^ Eichler, W. (1942). “Die Entfaltungsregel und andere Gesetzmäßigkeiten in den parasitogenetischen Beziehungen der Mallophagen und anderer ständiger Parasiten zu ihren Wirten” (PDF). Zoologischer Anzeiger. 136: 77–83. Bản gốc (PDF) lưu trữ ngày 4 tháng 3 năm 2017. Truy cập ngày 29 tháng 1 năm 2020.
- ^ Klassen, G. J. (1992). “Coevolution: a history of the macroevolutionary approach to studying host-parasite associations”. Journal of Parasitology. 78 (4): 573–587. doi:10.2307/3283532. JSTOR 3283532. PMID 1635016.
- ^ Vas, Z.; Csorba, G.; Rozsa, L. (2012). “Evolutionary co-variation of host and parasite diversity – the first test of Eichler's rule using parasitic lice (Insecta: Phthiraptera)” (PDF). Parasitology Research. 111: 393–401. doi:10.1007/s00436-012-2850-9. PMID 22350674.
- ^ Hafner, MS (1988). “Hafner MS, Nadler SA. Phylogenetic trees support the coevolution of parasites and their hosts. Nature. Mar”. pubmed.ncbi.nlm.nih.gov (bằng tiếng Anh). doi:10.1038/332258a0. PMID 3347269.
- ^ Brooks, Daniel R. (20 tháng 9 năm 1979). “Testing the Context and Extent of Host-Parasite Coevolution”. www.jstor.org (bằng tiếng Anh). tr. 299-307 (9 pages). doi:10.2307/2412584.
- ^ Rensch, B. (1950). “Die Abhängigkeit der relativen Sexualdifferenz von der Körpergrösse”. Bonner Zoologische Beiträge. 1: 58–69.
- ^ a b Dale, James; Dunn, Peter O.; Figuerola, Jordi; Lislevand, Terje; Székely, Tamás; Whittingham, Linda A. (7 tháng 12 năm 2007). “Sexual selection explains Rensch's rule of allometry for sexual size dimorphism”. Proceedings of the Royal Society of London B: Biological Sciences (bằng tiếng Anh). 274 (1628): 2971–2979. doi:10.1098/rspb.2007.1043. ISSN 0962-8452. PMC 2211517. PMID 17878139.
- ^ Blanckenhorn, Wolf; Dixon, A (2007). “Proximate Causes of Rensch's Rule: Does Sexual Size Dimorphism in Arthropods Result from Differences in Development Time?”. The American Naturalist. University of Zurich. 169 (2): 245–257. doi:10.1086/510597. PMID 17211807. S2CID 19831154.
- ^ Székely, Tamás; Freckleton, Robert P.; Reynolds, John D. (17 tháng 8 năm 2004). “Sexual selection explains Rensch's rule of size dimorphism in shorebirds”. Proceedings of the National Academy of Sciences of the United States of America (bằng tiếng Anh). 101 (33): 12224–12227. Bibcode:2004PNAS..10112224S. doi:10.1073/pnas.0404503101. ISSN 0027-8424. PMC 514460. PMID 15304645.
- ^ Lengkeek, W.; Didderen, K.; Côté, I. M.; van der Zee, E. M.; Snoek, R. C.; Reynolds, J. D. (1 tháng 10 năm 2008). “Plasticity in sexual size dimorphism and Rensch's rule in Mediterranean blennies (Blenniidae)”. Canadian Journal of Zoology. 86 (10): 1173–1178. doi:10.1139/z08-103. ISSN 0008-4301.
- ^ Fairbairn, D.J. (1997). “Allometry for Sexual Size Dimorphism: Pattern and Process in the Coevolution of Body Size in Males and Females”. Annu. Rev. Ecol. Syst. 28 (1): 659–687. doi:10.1146/annurev.ecolsys.28.1.659.
- ^ Piross, I.S.; Harnos, A.; Rozsa, L. (2019). “Rensch's rule in avian lice: contradictory allometric trends for sexual size dimorphism”. Scientific Reports. 9 (1): 7908. Bibcode:2019NatSR...9.7908P. doi:10.1038/s41598-019-44370-5. PMC 6536520. PMID 31133727.
- ^ Joel Asaph Allen (1877). “The Influence of Physical Conditions in the Genesis of Species”. line feed character trong
|title=tại ký tự số 37 (trợ giúp) - ^ “Allen's rule”.
- ^ Campbell và cộng sự: "Sinh học" - Nhà xuất bản Giáo dục, 2010.
- ^ "Sinh học 12" - Nhà xuất bản Giáo dục, 2019.
- ^ Lopez, Barry Holstun (1986). Arctic Dreams: Imagination and Desire in a Northern Landscape. Scribner. ISBN 0-684-18578-4.
- ^ Ashizawa, K. et al. (2007). Growth of height and leg length of children in Beijing and Xilinhot, China. In Anthropological Science. 116(1). Pages 67 & 68. Truy cập ngày 22 tháng 1 năm 2017, from link.
- ^ Gloger (1833)
- ^ Stresemann (1975)
- ^ Zink & Remsen (1986)
- ^ Dollo, Louis (1893). “Les lois de l'évolution” (PDF). Bull. Soc. Belge Geol. Pal. Hydr. VII: 164–166.
- ^ Gould, Stephen J. (1970). “Dollo on Dollo's law: irreversibility and the status of evolutionary laws”. Journal of the History of Biology. 3 (2): 189–212. doi:10.1007/BF00137351. PMID 11609651. S2CID 45642853.
- ^ Goldberg, Emma E.; Boris Igić (2008). “On phylogenetic tests of irreversible evolution”. Evolution. 62 (11): 2727–2741. doi:10.1111/j.1558-5646.2008.00505.x. PMID 18764918. S2CID 30703407.
- ^ Collin, Rachel; Maria Pia Miglietta (2008). “Reversing opinions on Dollo's Law”. Trends in Ecology & Evolution. 23 (11): 602–609. doi:10.1016/j.tree.2008.06.013. PMID 18814933.
- ^ “Luật Hebb là cơ sở tâm thần kinh học”.
- ^ Frost, Harold "New targets for fascial, ligament and tendon research: A perspective from the Utah paradigm of skeletal physiology" J Musculoskel Neuron Interact 2003; 3(3):201–209
- ^ Frost, Harold "The physiology of cartilagenous, fibrous, and bony tissue. C.C. Thomas, 1972
- ^ Frost, Harold "The physiology of cartilagenous, fibrous, and bony tissue. C.C. Thomas, 1972, page 176
- ^ Frost, Harold "Does the anterior cruciate have a modeling threshold? A case for the affirmative". J Musculoskel Neuron Interact 2001; 2(2):131–136
- ^ “Sinh Cơ Học Gãy Xương - Sinh học lành xương”. ortho.luanmd.com.
- ^ Anahad O'Connor (18 tháng 10 năm 2010). “The Claim: After Being Broken, Bones Can Become Even Stronger . Julius Wolff wrote his treatises on bone after images of bone sections were described by Culmann and von Meyer”. New York Times. Truy cập ngày 19 tháng 10 năm 2010.
This concept — that bone adapts to pressure, or a lack of it — is known as Wolff’s law. ... there is no evidence that a bone that breaks will heal to be stronger than it was before.
- ^ Frost, HM (1994). “Wolff's Law and bone's structural adaptations to mechanical usage: an overview for clinicians”. The Angle Orthodontist. 64 (3): 175–188. PMID 8060014.
- ^ Ruff, Christopher; Holt, Brigitte; Trinkaus, Erik (tháng 4 năm 2006). “Who's afraid of the big bad Wolff?: "Wolff's law" and bone functional adaptation”. American Journal of Physical Anthropology. 129 (4): 484–498. doi:10.1002/ajpa.20371. PMID 16425178.
- ^ Stedman's Medical Dictionary (Wayback Machine PDF)
- ^ Wolff J. "The Law of Bone Remodeling". Berlin Heidelberg New York: Springer, 1986 (translation of the German 1892 edition)